The Vital Question is a pretty wide-ranging book, touching on
- the alkaline vents hypothesis of abiogenesis (how life arose from non-life)
- what we can infer about endosymbiosis (how complex eukaryotic life arose from simpler bacteria and archaea – an incredibly unlikely event)
- the disposable soma theory (which this website’s chicken and egg logo represents)
- the mitochondrial-nuclear genome fit theory of longevity
This review is just about the first one, as it is the most clearly argued section of the book (and also the most interesting question). Endosymbiosis is a close second, but I find that I have no willpower to write a real review of that section. I wrote a romance short story about it; that’ll have to suffice.
Note: Lane chooses not to touch the major question of how DNA and DNA replication arose. This book is about how reality might have solved the other significant challenges.
Proposed timeline
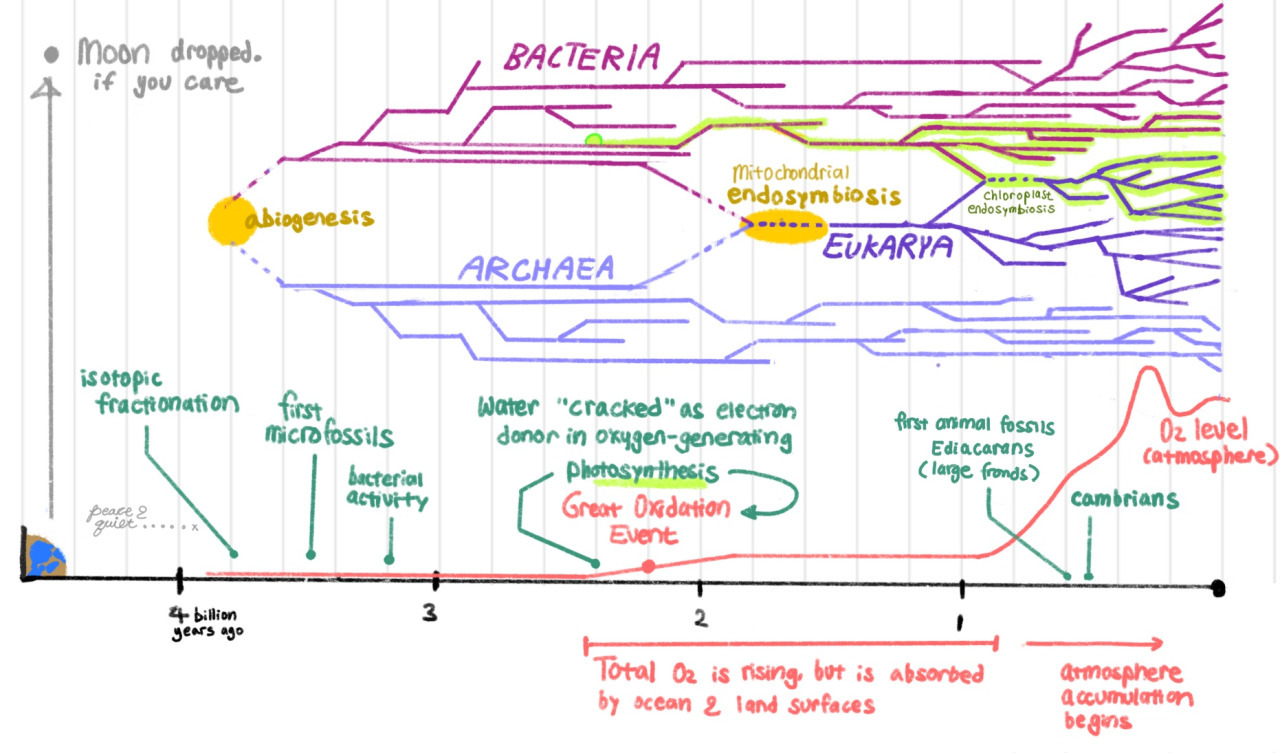
- 3.8bya: The earliest evidence for life is isotopic fractionation. The carbon (and iron, sulfur, nitrogen) atoms in graphite in Greenland are non-randomly sorted, which indicates the presence of cells whose enzymes have a slight preference for the lighter forms of each. However, geological processes can also produce non-random sorting, so this evidence is ambiguous.
- 3.5bya: Less ambiguously, we have microfossils that look like cells, again with isotopic signatures.
- We think bacteria and archaea split off really early, close to the beginning of life (abiogenesis) itself. This is because their cell walls and membranes are so different it’s hard to see how one could have evolved from the other. They probably emerged in parallel when they became independent at all.
- 3.2bya: We see bacterial activity in rust bands in rocks. When bacteria strip electrons from iron dissolved in the oceans, the oxidated iron precipitates out into rust and sinks down to the ocean floor.
- All bacteria and archaea respire (strip electrons from a donor ‘fuel’ to generate ATP, which fuels cellular activity). Many donors are possible – common ones are Fe2+, H2S, or H2O (which respectively become Fe3+, S, and O2 after the electron is stripped). Water is the last to be cracked as a donor.
- 2.4bya: It’s cracked now, by some bacteria (archaea never manage it), which leads to The Great Oxidation Event. The oxygen is first absorbed by the oceans, the seabed, and land surfaces. It’ll take over a billion years before the oxygen ‘sinks’ are exhausted, and oxygen accumulates in the atmosphere for real.
- 1.8bya: A bacterium somehow ends up inside an archaeon cell, and somehow they don’t die about it. The bacterium becomes a specialized energy-producing unit, allowing the host cell to grow larger and more complex. They become the first eukaryote, invent sex, and spawn all complex life.
- 1bya: This engulfment – endosymbiosis – happens one more time, to produce chloroplasts in plants.
- 0.5-6bya: Atmospheric oxygen levels are rising for real now. We see large complex eukaryotes for the first time. These creatures have specialized tissues whose failure threatens the entire organism, and it becomes advantageous to separate out the germline (the part of eukaryotes that divide forever, e.g. sperm and eggs) and make the rest of the body just durable enough to last as long as the most failure-prone tissues. Death by design has entered the world.
Abiogenesis
Nick Lane’s bet on the location of the origin of life is underwater alkaline hydrothermal vents, which are formed when water sinks underwater, reacts with certain common rocks like olivine, and comes back hot & alkaline & also holding a lot of rock stuff like metals and sulfur. When this re-meets the ocean, the rock stuff precipitates out into a “mineralized sponge” riddled with labyrinthine interconnected pores, throughout which the pH difference can be dramatically different. Like 3-5 pH units.
where again?
This rock offers a bit of natural inside/outside protection for a protocell putting itself together. Organics naturally concentrate in small spaces due to thermophoresis, which is the phenomenon where larger molecules accumulate in the colder regions of a small space (because they’re worse at randomly bouncing back to a hot region than a small molecule). So you have an environment full of CO2, hydrogen gas, metal catalysts, as well as a natural gentle flow of water carrying out waste. Promising!
Carbon fixation is the process of stripping carbon from inorganics like CO2 to tack them onto organic molecules like RNA. Every living cell has to do this. The simplest carbon fixation pathway only requires CO2 and H2, so let’s assume it’s the first pathway that was used. The pH difference comes into play twice, here: (1) Stripping an electron from H2 to give it to CO2 to make an organic molecule is hard in any pH, but not hard if the electron crosses a semiconducting mineral like FeS in a thin wall, from a more alkaline environment to a more acidic one. (2) Where does the energy to run these carbon fixation reactions come from? Simple membranes are by default porous to protons, which are small. Simple cells wedged between two areas with differences in pH can exploit the flow of protons, which go through (something like) ATP synthase to generate ATP, which can perform work in the cell much as they do now. (Actually, probably not ATP, but acetyl phosphate, which is much simpler but can do the same thing.)
But howww do you go from passively exploiting the proton gradient to producing it yourself by pumping protons out, as modern cells do? As a hint, we notice that all cells seem adapted to an internal concentration of Na+ that’s weirdly lower than the ocean concentration. Na+ can also be used for energy production, and lipid membranes are much less permeable to Na+ than H+. So while there’s no point in pumping H+ that’ll come back immediately, there is an advantage to pumping Na+, since it’ll stay out and increase the energy gradient you’re using to live. The additional Na+ gradient can give the cell 60% more power than relying on protons alone, meaning cells with the Na+ pump can colonize areas of the vent with a smaller pH difference. Once you have an Na+ pump, there’s an advantage to tightening the membrane against protons since… I didn’t believe the logic here :/ but the endpoint is that you have an impermeable membrane studded all over with proton pumps, at which point you’re ready to leave the vents.
(Even this relatively plausible account is so implausible I kind of don’t believe life actually arose.)
Archaea vs Bacteria split
Archaea and bacteria probably diverged before attaining independence, and left the vents separately. We think this because their cell membranes and cell walls are so different. Like, they both use glycerol to make their membrane impermeable to protons, but they use different stereoisomers of glycerol to do so. Both of them use the same fundamental energy converting membrane protein, but oriented in opposite directions, which led to different design constraints.
(Which I do not understand well enough to relay to you, even after reading the relevant section 4-5 times.)
They do both have ATP synthase, from which we can infer the order of developments in very early life:
- DNA and DNA replication arises. So does using ATP for energy, and ATP synthase to continually rebuild ATP.
- One strain of proto-cell starts tightening its membrane against protons using glycerol, colonizes the shallower-pH-gradient parts of the vent, and eventually leaves the vent.
- Some time later, a second strain independently does the same, using a different handedness of glycerol, etc.
These two branches coexist and probably swap DNA for over a billion years, but when an archaeon somehow engulfs a bacterium, the bacterium becomes mitochondria and the merger spawns all of complex eukaryotic life.
Endosymbiosis happens again several times, most dramatically when a photosynthetic bacterium is engulfed by eukaryotic life and the merger results in plants, but as far as we know, de novo endosymbiosis only happened once for certain.